|
|
Главная |
|
|
Пишите |
|
Оглавление
Эволюция генетического кода видна в самом коде — он как бы "застыл" в своём развитии. Об этом говорит и его избыточность. На основе знаний о первичных аминокислотах можно предложить гипотетическую модель его дуплетного протокода.
Дуплетный код короче нашего триплетного на треть. Соответственно, меньше будет и поперечный размер протоклетки. Возможно, именно нанобактериии и ультрамикробактерии — носители этого пракода. Их размеры не соответствуют риплетной ДНК, но вполне подходят под диплетную РНК.
Нанобактерии — одно из самых нижних звеньев пищевой цепи на Земле. Об этом косвенно свидетельствует и явление литофагии — поедания глины в некоторых обществах.
Формами, родственным нанобактериям, могут быть похожие структуры в марсианском грунте, в марсианском метеорите, обнаруженном в Антарктиде, а также в "кровавых дождях", последние известные события которого произошли в Индии в прошлом веке. Не исключено, что жизнь на Землю перенесена с Марса, где ее эволюция замерла на протоклетках с дуплетным прагенетическом коде.
В этом исследовании описывается дуплетная модель протокода и излагаются аргументы в пользу его современных носителей — нанобактерий.
Ключевые слова: эволюция генетического кода, прагенетическая кодировка, носители нуклеотидного пракода, нанобактерии, глинобактерии, природа литофагии, поедание глины, кровавый дождь в Индии, бактерии марсианского метеорита.
Keywords: genetic code evolution, proto-genetical encoding, nanobacteria, nucleotide proto-code carriers, lithophagia, bloody rain in India, bacteria in Martian meteorite, halobacteria, clay eating.
Генетический код (ГК) — единая для земной биоты система кодирования и хранения наследственной информации в молекулах нуклеиновых кислот в виде последовательности 3-х нуклеотидов из 4-х возможных (рис. 1). Нуклеотиды отличаются азотистыми основаниями: и обозначаются буквами А (аденин), Т (тимин), Г (гуанин), Ц (цитозин). Т.о., общее число триплетов (кодонов) — 43 = 64. Всего аминокислот — 20, т.е., код избыточен (вырожден).
Во всех ли ныне живущих видах ГК одинаков? Един ли он в древнейших остатках земных организмов? Универсален ли он во Вселенной? Несёт ли он следы эволюции и развивается ли дальше? На некоторые вопросы наука уже ответила. На некоторые в обозримом будущем сложно будет ответить. А в этой статье попробуем, анализируя ГК, чуть заглянуть в его прошлое. Параллельно, как увидим, будут получены ответы на много других связанных вопросов (см. Заключение).
Проведём анализ:
Проведём анализ в соответствии с составом предмета исследований (гл. 3).
Генетический код удобно представить в виде следующей таблицы:
| 1-е осн. | Второе основание | 3-е осн. | |||||||
| Гуанин | Урацил | Аденин | Цитозин | ||||||
| Ц | Арг** |
NH2
/
R-(CH2)3-NH-C
\
NH
|
Лей* |
CH3
/
R-CH2-CH
\
CH3
|
Гис** |
R-CH2-C=CH-N
\ ||
NH-CH
|
Про |
R-CH2 | | H2C-CH2 |
У |
| Арг** | Лей* | Гис** | Про | Ц | |||||
| Арг** | Лей* | Глн | R-(CH2)2-CONH2 | Про | А | ||||
| Арг** | Лей* | Глн | Про | Г | |||||
| Г | Гли | R-H | Вал* |
CH3
/
R-CH
\
CH3
|
Асп | R-CH2-COOH | Ала | R-CH3 | У |
| Гли | Вал* | Асп | Ала | Ц | |||||
| Гли | Вал* | Глу | R-(CH2)2-COOH | Ала | А | ||||
| Гли | start (был Вал?) | контрольный | Глу | Ала | Г | ||||
| А | Сер 2 (был start?) | R-CH2-OH | Изо* |
CH3
/
R-CH
\
CH2-CH3
|
Асн | R-CH2-CONH2 | Тре* |
OH
|
R-CH-CH3
|
У |
| Сер 2 (был start?) | Изо* | Асн | Тре* | Ц | |||||
| Арг** 2 (был stop?) |
NH2
R-(CH2)3-NH-C<
NH
|
Изо* | Лиз* | R-(CH2)4-NH2 | Тре* | А | |||
| Арг** 2 (был stop?) | Мет* (был Изо?) | R-(CH2)2-S-CH3 | Лиз* | Тре* | Г | ||||
| У | Цис | R-CH2-SH | Фен* | R-CH2-B | Тир | R-CH2-B-OH | Сер | R-CH2-OH | У |
| Цис | Фен* | Тир | Сер | Ц | |||||
| stop (был Три?) | контрольный | Лей* 2 (был Фен*?) | R-CH2-B/2 | stop (был Тир/Мет?) | контрольный | Сер | А | ||
| Три* |
NH
R-CH2-C2H < |
B
|
Лей* 2 (был Фен*?) | stop (был Мет?) | Сер | Г | ||||
Примечания и обозначения:
Сокращения для аминокислот (в скобках - количество их кодонов) и их "вес" (кол-во атомов кроме радикала и водорода):
|
|
Из них незаменимых — только половина (10).
Расположим аминокислоты по весу (с бензольным кольцом — отдельно ниже: B=6):
|
|
Первые 10 можно считать простыми (Гли, Ала, Сер, Асп, Цис, Асн, Вал, Глу, Про, Тре), другие 10 — сложными. Всего же для дуплетного кода наберется 15 аминокислот с весом меньше 6. Что интересно, изпростых аминокислот — незаменимых всего 2 (валин и треонин).
Какие видим закономерности в столбцах и строках генетического кода:
Что можно еще увидеть и предположить при исследовании генетической кодировки:
Следовательно, можно предположить следующие цепочки формирования одних аминокислот из других:
Какие ещё замечания возникают при анализе аминокодировки:
С учетом предыдущих замечаний, сначала кодируемых аминокислот было меньше (также без фенилаланина, цистеина, глутамина и глутаминовой кислоты) — 20 − 6 = 14. Кстати, указанные отсутствующие аминокислоты и по структуре сложнее.
Изначально, вероятно, код был "2-буквенный" (дуплетный), а первичных кислот могло быть не больше 4 x 4 = 16. Можно предположить следующую схему их кодирования:
| \ | Гуанин | Урацил | Аденин | Цитозин |
| Цитозин | Арг** | Лей* | Гис**/Глн? | Про |
| Гуанин | Гли | Вал* | Асп/Глу? | Ала |
| Аденин | Сер/Арг**? | Изо* | Асн?/Лиз* | Тре* |
| Урацил | Цис/Три*? | Фен* | Тир | Сер |
(В вышеприведенной таблице нет места для метионина*)
Возможно, в клетке "Сер/Арг" располагался аспарагин (как более похожий на серонин), и тогда в клетке "Асп/Глу" находился только глутамин.
Место Лей-2 мог занимать фенилаланин — тогда среди древнейших (первичных, двузнаковых) аминокислот была одна с бензольным кольцом (интересно, что это даёт?). А, может быть, место для Лей-2, на самом деле было резервным или отдано командным кодам.
Если в спорных местах выбрать именно незаменимые аминокислоты, то эта табличка будет такой:
| \ | Гуанин | Урацил | Аденин | Цитозин |
| Цитозин | Арг** | Лей* | Гис**/Глн? | Про |
| Гуанин | Гли | Вал* | Асп/Глу? | Ала |
| Аденин | Арг**? | Изо* | Лиз* | Тре* |
| Урацил | Три*? | Фен* | Тир | Сер |
(В вышеприведенной таблице отсутствуют метионин и цистеин)
А если в спорных местах выбрать более простую, то табличка будет такой:
| \ | Гуанин | Урацил | Аденин | Цитозин |
| Цитозин | Арг** | Лей* | Глн | Про |
| Гуанин | Гли | Вал* | Асп | Ала |
| Аденин | Сер | Изо* | Асн | Тре* |
| Урацил | Цис | Фен* | Тир | Сер |
(В вышеприведенной таблице отсутствуют гистидин, глутамин, лизин, метионин)
Самые малые по весу — Гли, Ала, Сер, Асп, Цис, Асн, Вал, Глу, Про, Тре. Они здесь все, кроме Глу. Может быть, глутаминовая кислота занимала один из кодонов, где серонин?
Незаменимыми для взрослого здорового человека являются 8 аминокислот: валин, изолейцин, лейцин, лизин, метионин, треонин, триптофан и фенилалани́н. Для детей незаменимыми также являются аргинин и гистидин. В таблице представлены почти все они, кроме метионина и фенилалани́на — код последнего, возможно, соответствовал ячейке с Лей 2.
Расcмотрим теперь эти выводы с коэволюционной позиции (источник: http://scorcher.ru/theory_publisher/show_art.php?id=216). В 1975 г. Вонг предположил, что главным фактором совместной эволюции генетического кода и аминокислот являлись их биосинтетические свойства [Wong J.T.F. 1975, 1981]. По его мнению, современные ферментативные пути биосинтеза аминокислот должны в значительной степени быть сходны с путями синтеза, возможно в том числе и абиогенного на стадии формирования кодируемого пептидного синтеза. Такая сложная система, как ГК не могла возникнуть сразу полностью. Первоначально, вероятно, в кодируемый синтез были вовлечены лишь несколько аминокислот.
Основываясь на современных знаниях о биосинтезе аминокислот [Cohen G.N. 1968; Rodwell V.W. 1969] Вонг предложил следующие пары, связанные отношением «предшественник-продукт»:
|
|
|
Сокращаем эту цепочку до 6 первичных аминокислот (не считая самого простейшего глицина), на основе которых синтезируются 13 других:
Исследовав функции названных поздних (вторичных, триплетных) аминокислот, можно определить, каких процессов не было в организмах, имеющих этот дуплетный генетический механизм. Поскольку на Земле не обнаружено их ни живыми ни ископаемыми, можно предположить, что сначала жизнь развивалась вне Земли.
Согласно Г. Хартману (Hartman, 1995), вначале были простые синглетные, затем дуплетные соответствия нуклеиновых кислот аминокислотам. При этом GC-пapa нуклеотидов могла кодировать наиболее простые и распространенные аминокислоты: GG кодирует глицин, GC — аланин, СС — пролин, CG — аргинин. Считается, что в начале в построении полипептидов принимали участие не более 10 аминокислот (Juke, 1973). Е.Трифонов (Trifonov, 2000) предположил, что первичным был триплет CGU и его ближайшие производные. Он оценил статистически, какие из 20 аминокислот с наиболее высокой вероятностью могли принять участие в начальной эволюции генетического кода. Оказалось, что эти вероятности образуют ряд: Гли, Ала, Вал, Асп, Глу, Про, Сер, Лей, Тре, — почти точно совпадающий относительной распространенности аминокислот, полученной в экспериментах по их абиогенному синтезу [Галимов 2001, с. 120].
У клеток с дуплетным генетическим кодом и ДНК (или, скорее вмего, РНК) должна быть минимум на 1/3 короче, и сами они меньше. Возможно, эти протоорганизмы были размером с нанобактерий, составляющих, между прочим, большую часть земной биомассы (как "тёмная материя" имеет большую массу, чем обычная). Более того, вполне вероятно, что сами нанобактерии являются "двухбуквенными системами". Ведь до сих пор из них не извлечено ДНК, которое "определяется" лишь по косвенным признакам. Значит и генетический механизм нанобактерий неизвестен. А, т.к. подобные органические включения (сферы и цепочки) найдены в "марсианских метеоритах", то, не исключено, жизнь возникла (или сначала существовала) до Земли на Марсе.
Малые размеры нанобактерий можно объяснить тем, что генетический механизм их не 3-буквенный, а 2-буквенный — тогда и ДНК (вернее, РНК) будет на треть короче. Если так, то, предположительно, на треть меньше объёма будут занимать и сами эти 2-буквеннонаследуемые организмы (возьмите 2 веревки равной толщины, но одна короче другой на ⅓, и скрутите их в комочки — очевидно, и объём того комочка будет меньше на 1/3). Значит, размер таких гипотетических "бикодобактерий" будет в диаметре еще меньше.
Если убрать из ДНК клетки треть кода (Ln = ⅔La), получим относительный размер дуплетно-кодируемой клетки на основе следующих вычислений:
где "a" - индекс прокариот (например, археобактерий), а "n" - "2-буквенных" (дуплетногенных) бактерий (условных нанобактерий); L - длина ДНК; R, D и V - радиус, диаметр и объём клетки.
Считается , что минимально допустимый размер живой сферы (с триплетным ДНК) – 0,15—0,20 мкм. Значит, минимальный размер «дуплетной» клектки будет 0,15 × 0,875 ~ 0,13. А если генетической молекулой является однонитевая РНК, как в ретровирусах [Вирусология 1989, с. 434], то наименьший размер снизится до примерно 0,06 мкм. Размеры же обнаруженных наноклеток составляют 0,05-0,2 мкм [Воробьева 2007, с. 383], что вполне сопоставимо с полученным результатом.
Из этого делаем вывод, что нанобактерии, возможно, являются организмами с дуплетным кодированием генетической информации (причём, не исключая, что на основе однонитиевой РНК), и они – живые, а не комочки каталитической глины.
К сказанному добавим, что в [Adams 2001] определено, что наноклетки с диаметром 50 нм могли содержать 8 генов и обладать способностью реплицировать свою ДНК и синтезировать белки; генерировать и сохранять энергию; осуществлять активный и пассивный транспорт веществ через мембрану — клеточные функции генного класса.
Официальной датой открытия каменной бактерии, названной за ее малые размеры нанобактерией, считается 1990 год. Впервые ее обнаружил американский геолог Техасского Университета Роберт Фольк. Используя раструбную электронную микроскопию, он выявил колонии размножающейся карликовой бактерии, покрытой каменной оболочкой среди горных пород, вулканического туфа, арагонита, доломита.
Нанобактерии первоначально описаны как карликовая форма бактерии, преимущественно достигающая 0,05—0,2 мкм (50-200 нанометров) в диаметре, т.е. сопоставима или даже меньше, чем некоторые вирусы. Для справки: размеры вирусов составляют 0,1—1,0 мкм: например, у безоболочечных аденовирусов 70—90 нм, у иридовирусов 125—300 нм, у крупных поксвирусов (300—450) × (170—260) [Вирусология 1989, сс. 27-30]. Имеются также микробактерии с кольцевой хромосомой (риккетсии) размерами 0,2—2,0 мкм (чуть больше нанобактерии). Вообще, размеры прокариотных клеток в большинстве случаев колеблются от 0,5 до 3 мкм, а самые мелкие (микоплазмы — клетки без стенки) не превышают 0,10–0,15 мкм (фактически, сопоставимы с нанобактериями).
Возможно, большая часть биомассы Земли состоит именно из нанобактерий, которые участвуют в мировом круговороте минералов и органики. Они найдены в древнейших (докембрийских) породах Земли, а похожие структуры найдены в антарктическом метеорите ALH 84 001, прилетевшим с Марса 13000 лет назад.
Сами глины в измененных земных вулканических скалах могут поглощать и полимеризовывать аминокислоты в длинные цепочки полипептидов, считающихся основными "строительными блоками" жизни. Близкая ассоциация между этими каталитическими глинами и потенциально наиболее примитивной формой жизни, какой являются нанобактерии, может способствовать пониманию причин возникновения жизни на Земле.
Живая природа нанобактерий до сих пор признается не всеми — нанобактерии могут быть "всего лишь" самоорганизующимися частичками карбоната кальция (мела). Некоторые рассматривают их не как биологические, а как органические структуры, т.к. ДНК обнаружены по косвенным признакам (окрашивание), а реально не извлечены, и размер меньше минимума (140 нм).
Другие, напротив, считают их возбудителями мочекаменной болезни, туберкулеза, атеросклероза [т.е., всё, что связано с отложением кальция, солей и камней, а следовательно, и артроза] и др., даже рака. В "бессмертных организмах" (один из видов медуз) их не найдено — из этого делают вывод, что нанобактерии могут быть причиной старения.
Нанобактерии содержат: белок, липиды, ДНК, рибонукленовую кислоту, АТФ, аминокислоты. По данным финских исследователей, в химический состав нанобактерии входят: неорганического фосфата — 39%, N — 1—13%, Р — 12,3—14,6%, кальций — 23,4—23,5%, Mg — 1,4—1,9%; К — 0,1%, Na — 1,2—1,42%. Kajander А.О. et ai приводят доказательство способности нанобактерий осаждать кальцит, арагонит, фосфаты [их первый признак жизни?]. Возможно, нанобактсрия использует их как строительный минерал своей каменной мантии.
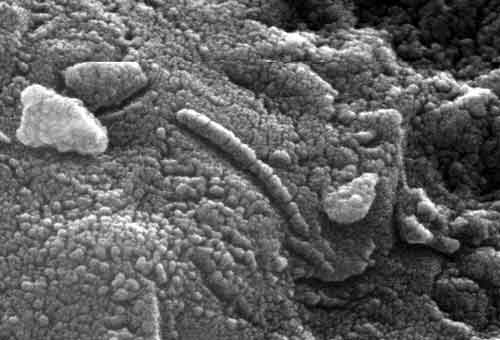
Что касается упомянутого марсианского метеорита ALH 84 001, обнаруженного в Антарктиде, то объем обнаруженных в нём организмообразных структур в тысячу раз меньше самых мелких земных бактерий. Одним из доводов, что это не следы живой органики, является утверждение, что кристаллы образовались при +250℃. Но недавно в "кровавом водопаде" на Антарктиде обнаружили микроорганизмы, которые размножаются при +315°C! Небольшой камень весом около 2 кг был обнаружен экспедицией по поиску метеоритов в антарктическом районе Аллан Хиллз. Находку назвали ALH84001 (ALH расшифровывается как Allan Hills). Исследовав состав пород метеорита, ученые заключили, что он попал на Землю с Марса. После изучения изотопного состава некоторых элементов, входящих в состав метеорита, специалисты сделали вывод, что порода, из которой "родился" ALH84001, сформировалась на Красной планете приблизительно 4,5 млрд. лет назад. Это очень солидный возраст, особенно если принять во внимание, что Марс как таковой образовался всего на 100 миллионов лет раньше. |
|
Рис. 1. Организмообразные структуры в грунте Марса. |
Где-то в промежутке от 3,6 до 4 млрд. лет назад участок породы, который позже станет ALH84001, был поврежден в результате падения метеорита. Поверхность покрылась трещинами, которые со временем заполнились водой. В лужицах накапливались соединения углерода. Примерно 16 миллионов лет назад фрагмент породы, еще не имеющий названия, откололся в результате удара другого метеорита. Следующие 15 миллионов 967 тысяч лет кусок марсианской скалы провел в космическом пространстве (время пребывания ALH84001 в космосе ученые вычислили, оценив степень повреждения его поверхности космическими лучами). Наконец, 13 тысяч лет назад камень упал на Землю, где пролежал вплоть до 27 декабря 1984 года.
Цито-подобные структуры найдены и в марсианском грунте в ходе исследования НАСА (рис. 1).

Красные (с железом) глины на Марсе (рис. 2) - возможно, продукт деятельнолсти нанобактерий. По данным Р. Фолька железо является одной из сред обитания нанобактерий, которые найдены им в продуктах коррозии железа. Получена высокая степень корреляции между уровнем нанобактерий в 1 мл питьевой воды и концентрацией суммарного железа (г=+03933) [Волкова 2007, с. 99]. |
|
Рис. 2. Красные глины Марса. |
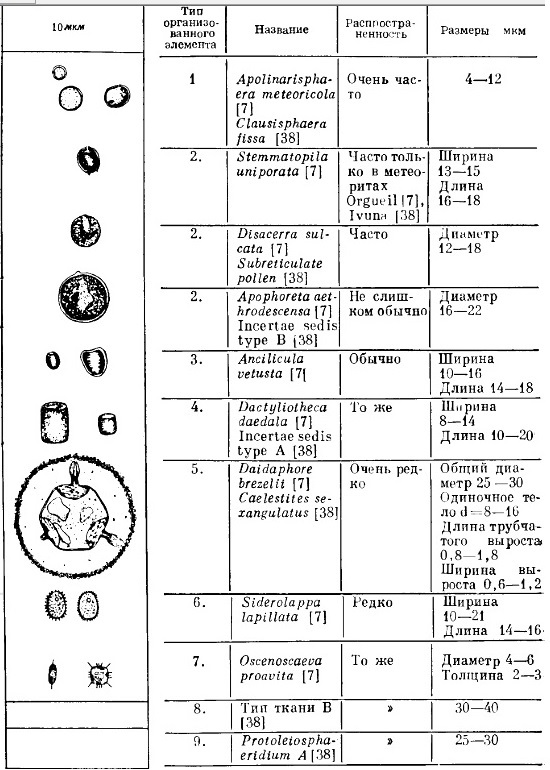
Так называемые "организованные элементы" [Руттен 1973, с. 370], обнаруженные в метеоритах особого типа, называемых углистыми хондритами, показывают внешнюю и масштабную схожесть с элементами земной биоты — нанобактериями, икосаэдрическими вирусами, пыльцой (рис. 3) и др. Размеры этих организованных элементов — от 4 до 30 мкм, что больше обычного размера бактерии (0,5—3 мкм). |
|
Рис. 3. Организованные элементы в углистых хондритах. |
Возможно, именно нанобактериями обусловлены известные "кровавые дожди" в Индии.

В индийском штате Керала в 2001 году прошёл кроваво-красный дождь. Он лился 2 месяца. В 2012 году кровавый дождь выпадал и на Шри-Ланке. Лабораторные исследования показали, что дождь окрасился в такой цвет из-за спор лишайника Trentepohlia annulata, которые в большом количестве выделились во влажный воздух в сезон дождей. Эти споры содержат много каротиноидов - изопреноидных углеводородов общей формулой C40H56. Цветные осадки - хоть и редкое, но не небывалое явление. Красные дожди могут возникать, например, в зонах повышенной кислотности или из-за пылевых бурь. В 2006 году в Колорадо выпадал розовый снег, очевидцы утверждают, что он пах арбузом. В феврале 2015 года в Саратовской области выпадал оранжевый снег: оказалось, что циклон, пришедший из Северной Африки, притащил с собой частицы песка из-за пустыни. |
Получается, вся жизнь произошла из глины — как и сказано в Писании о создании человека. В [Паничев 2011] системно рассматриваются факты и причины поедания глины человеком и животными (феномен литофагии). Возможно, это говорит о том, что глина — самое нижнее звено пищеварительной цепи в земной биосфере, которое, вероятно, включает большую нанобактериальную массу.
Похожий феномен — геофагия — наблюдается в поведении летучих мышей и других рукокрылых, которые поедают грязь и вылизывают камни, чтобы обеспечить противоядие от растительных ядов, которые случайно попадают к ним после употребления в пищу фруктов.
Не исключено, что старение связано как раз с "закоксовыванием", вернее, "оглиниванием" организма этими "глинотельцами". Т.о., поговорка, что к старости человека тянет к земле, увы, имеет реальную причину.
Земная кора материков состоит из гранитов, а океаническая — из базальтов. Базальты есть на всех планетах земной группы (и на Луне), а гранитная — только на Земле, и немного найдено на Марсе. На Венере тоже найдены похожие на гранит минералы - гнейсы, но там меньшее количество кремнеземов. Континентальная кора возникла 3,8 миллиардов лет назад синхронно с первыми обнаруженными признаками жизни на планете.
Основной состав гранитов — SiO2 и Al2O3 — такой же, как в глине. Если глина — продукт жизнедеятельности нанобактерий (которых не даром называют еще "глинобактериями"), то логично предположить, что материковую кору Земного шара образовала именно эта нанобиота, так же как незначительный её слой на Марсе.
Наличие гнейсов на Венере может говорить о том, что они сформировались из переплавки лёгких материалов венерианской литосферы с включением околоорганических веществ из планетоземалей, остатками (или представителями) которых могут являться углистые хондриты — наиболее распространённая подгруппа метеоритов, содержащие хондры — сферические или эллиптические образования преимущественно силикатного состава. Они появились уже 4,5 млрд. лет назад. Затем, при развитии биосферы на планете, осадочные глино-органические материалы после переплавки становятся гранитами и формируют материковую кору планеты.
Сравнительная таблица реальных и предполагаемых бактерий и добактериальных форм:
| № | Размер (мкм) | Название микробиообъекта | Место обнаружения | Среда жизнедеятельности |
| 1. | ||||
| 2. | ||||
| 3. | ||||
| 4. | ||||
| 5. | ||||
| 6. | ||||
| 7. | ||||
| 8. | ||||
| 9. | ||||
| 10. | ||||
| 11. |
В данном исследовании получены следующие результаты:
На основании этих результатов приходим к выводам:
Здесь приведены некоторые термины и аббревиатуры, встречающиеся в статье.
Термины, принятые в науке, тем не менее объясняются и в самой статье при их первом применении, например, геофагия, глинобактерии, дуплетный или триплетный (о ДНК и генетическом коде) в т.ч. дуплетногенный, литофагия . Используются также принятые однобуквенные сокращения для азотистых оснований (как самые частые в употреблении): А (аденин), Т (тимин), Г (гуанин), Ц (цитозин).
Единицы измерения в тексте статьи не разъясняются, например, нм - нанометр (10-9 метра).
Часть терминов и неологизмов придумана и предложена автором для возможного будущего применения (и представляют собой взгляд на явления под новым углом): глинотельцы, нанобиота, наноклетка (не очень корректное название), цито-структуры (в см. клеточные).
Впрочем, что-то мог напутать между этими двумя частями.
В основном здесь пока библиография трудов по биологическим тематикам: вирусологии, биогенезе, генетике, микробиологии, палеонтологии...
Упоминаемые в статье сетевые источники информации и другме полезные веб-ссылки на тему эволюции генетического механизма.
Доклад по данной теме впервые был зачитан в октябре 2017 года на XII Международном конгрессе "Космос и биосфера" в Алуште (Крым).
Все внутренние ссылки в этой статье (не указанные в разделе литературы и не ведущие к отдельному разделу статьи), ведут на различные страницы авторского сайта garshin.ru, информация на которых имеет разную степень неточности, служа исключительно как "сырьё" для дальнейшего поиска точных сведений, а также как "пища" к размышлению и набор авторских гипотез.
Такими страницами-полигонами ("черновиками") являются, в первую очередь, следующие:
Другие страницы моего сайта (приведены по алфавиту), где приводятся материалы и исследуются темы, затронутые в статье:
Ключевые слова для поиска сведений по анализу кодировки генов:
На русском языке: эволюция генетического кода, прагенетическая кодировка, носители нуклеотидного пракода,
глинобактерии, нанобактерии, организмообразные структуры, бактерии марсианского метеорита, кровавый дождь в Индии,
причина геофагии, природа литофагии, поедание глины;
На английском языке: genetic code evolution, proto-genetical encoding, nanobacteria, nucleotide proto-code carriers,
lithophagia, bloody rain in India, bacteria in Martian meteorite.
|
|
|
|
|
|