|
|
Главная |
|
|
Пишите |
|

Период определен только хронометрически, геологический слой земных пород не выделяется [тогда это фикция?]. Ко времени неоархея относится Беломорский цикл, в который происходило формирование настоящей континентальной земной коры. Кислородный фотосинтез впервые появился в этой эре, в результате чего концентрация кислорода в атмосфере Земли достигла уровня Пастера. Числа (точки) Л. Пастера - критические для эволюции биосферы уровни содержания кислорода в атмосфере Земли. Различают основное и дополнительное числа Л.Пастера; оба уровня достигнуты в архее. 1. Основное число Л.Пастера - достижение содержания кислорода в атмосфере в размере 1% от современного уровня, что привело к возможности возникновения аэробов [сначала в виде эубактерий?]. 2. Дополнительное число Л.Пастера - достижение содержания кислорода в атмосфере в размере 1О% от современного; момент формирования озоносферы, что позволило организмам заселить мелководья, а затем выйти на сушу. В конце неоархея (на границе с палеопротерозоем 2,5 миллиардов лет назад) в Австралии при исследовании железистых формаций в районе Пилбара найден пласт с ударными сферулами, обычно являющиеся признаком падения на планету метеорита. |
Разделы страницы о неоархейскской эре:
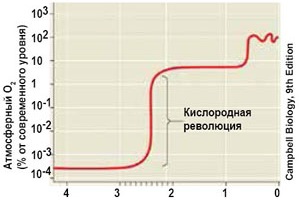
В истории атмосферного кислорода имеют значение несколько его пороговых величин. На Земле, лишенной фотосинтеза, кислород образуется в атмосфере за счет фотодиссоциации молекул воды. Его содержание, по расчетам Г. Юри, не может превышать 0,001 (0,1%) от современного (точка Юри) и автоматически держится на этом уровне. При таком содержании кислорода может существовать только анаэробная жизнь. Появление молекулярного кислорода за счет фотосинтеза сделало возможным для живых клеток процесс дыхания, который представляет намного более эффективный путь высвобождения энергии, чем анаэробное брожение. С этих позиций важна величина 0,01 (1%) содержания кислорода от современного уровня – так называемая точка Пастера. Существует целый ряд микроорганизмов, способных переключать свой энергетический обмен с дыхания на брожение и обратно при колебаниях кислорода ниже или выше точки Пастера. Есть предположения, что в раннем и среднем протерозое подобное переключение долгое время служило регулятором содержания О2, так как при повышении его концентрации кислород начинал интенсивно расходоваться на дыхание. Вместе с тем жизнь получила возможность распространиться почти до поверхности водоемов, так как ультрафиолетовые лучи за счет слабого озонового экрана могли теперь проникать на глубины не более метра. Третье пороговое содержание О2 (точка Беркнера-Маршалла) [или дополнительное число Пастера] соответствует 10 % от современного. Оно определяет такую сформированность озонового экрана, при которой потоки жестких ультрафиолетовых солнечных лучей уже не достигают земной поверхности и не препятствуют развитию жизни. По расчетам 60-х годов, точка Пастера была перейдена менее 1 млрд лет назад, а точка Беркнера-Маршалла – к концу силура, что обеспечило возможность выхода жизни на сушу. Современные исследования заставляют предполагать гораздо более раннее развитие событий. По некоторым данным, переход точки Пастера мог произойти уже 2,5 млрд лет назад (рубеж архея и протерозоя), а 10-процентное содержание кислорода было достигнуто уже в период 1,8–2,0 млрд лет от современности (орозирий). Ещё одна новость. Внутри полостей канадского кварца обнаружены микроскопические капли нефти, которая могла сформироваться только при наличии кислорода в атмосфере. Т.к. возраст минерала 2,5 миллиардов лет, то дата насыщения земной атмосферы кислородом должна быть отодвинута почти на 500 миллионов лет в прошлое, т.е. к 3,0 Ga (гигалет назад), а это - середина мезоархея. А отпечатки дождевых капель на породах (вулканическом пепле), возраст которых оценивается в 2,7 млрд лет, помогли определить состав атмосферы и условия на тогдашней Земле. Причём, глубина от капель в песке свидетельствует об их скорости (а также тогдашней силе тяжести), что позволяет определить плотность воздуха. В то время Земля вращалась вокруг своей оси гораздо быстрее, Луна была гораздо ближе, а солнечное излучение - гораздо слабее. 2,7 млрд лет назад на Земле еще не было [эукариотической] флоры и фауны, а атмосфера была непригодна для дыхания. Азота было немало, как и сейчас, но кислорода не было совсем. Скорее всего, вместо кислорода в атмосфере присутствовали такие парниковые газы, как метан и углекислый газ. Благодаря свойствам этих газов удерживать тепло, Земля как бы была укутана согревающим одеялом [Солнце-то ещё не сильно пекло]. Таким образом, на протяжении более двух миллиардов лет биосфера формировалась исключительно деятельностью прокариотов. Они полностью изменили геохимическую обстановку на Земле: сформировали кислородную атмосферу, очистили ее от токсических вулканических газов, связали и перевели в карбонатные породы огромное количество СO2, изменили солевой состав океана и сформировали громадные месторождения железных руд, фосфоритов и других ископаемых. Формирование окислительной атмосферы повлекло за собой бурное развитие эукариотической жизни, энергетика которой основана на процессе дыхания. Очевидно, что эукариотная жизнь тесно связана с аэробной средой, подготовленной для нее прокариотами. Первые аэробные организмы могли возникнуть довольно рано в составе цианобактериальных сообществ, которые, по выражению палеонтологов, были своего рода "кислородными оазисами" в анаэробной среде. |
Можно предположить, что появление эукариот было приурочено к такому периоду истории биосферы, когда условия были особенно нестабильны и непредсказуемы; когда приспособительная стратегия прокариот (быстрое мутирование, горизонтальный обмен генами и отбор устойчивых клонов) оказалась слишком расточительной и недостаточно эффективной. Именно в такой ситуации большое преимущество могла получить принципиально более универсальная и экономичная адаптивная стратегия, основанная на развитии адекватной (целесообразной) модификационной изменчивости.
Это предположение косвенно подтверждается палеонтологическими и геологическими данными. Древнейшие достоверные следы присутствия эукариот (углеводородные биомаркеры – холестан его 28-и и 30-и углеродные аналоги) обнаружены в отложениях возрастом 2,7 * 109 лет, т.е. в верхах Архея (Brocks et al., 1999). Именно к этому времени, считается, приурочено начало эпохи самых грандиозных за всю историю Земли геологических преобразований.
Согласно О.Г.Сорохтину и С.А.Ушакову (2002), на рубеже Архея и Протерозоя произошло формирование железного ядра Земли [оно наполовину было сформировано даже раньше - не позже 3,2 * 109 лет назад], что привело к целому комплексу быстрых и даже катастрофических перемен в литосфере, гидросфере и атмосфере. Сильнейшие конвективные течения в мантии привели к образованию единого суперконтинента – "Моногеи" (Кенорленда: 2,7-2,1 гигалет назад); на этот период приходится пик тектонической активности.
Чуть позже, в раннем протерозое, "после начала гидратации ультраосновных пород и образования серпентинитового слоя океанической коры произошло связывание в карбонатах плотной углекислотной атмосферы архея, исчезновение парникового эффекта и как следствие этого резкое похолодание климата" (Сорохтин, Ушаков, 2002, с. 283). На этом же рубеже произошло резкое повышение pH морской воды (от 3-5 в конце Архея до 7-8 в начале Протерозоя).
Эти катастрофические события могли способствовать появлению и экспансии первых эукариот двумя путями:
В частности, сильнейшеее похолодание и последовавшее за ним Гуронское оледенение (2,4—2,0 млрд. лет назад
как часть Канадской криоэры)
должно было крайне негативно сказаться на теплолюбивых цианобактериальных сообществах.
Не менее важным негативным фактором должно было стать предполагаемое катастрофическое снижение концентрации углекислого газа,
что для фотосинтетиков означает голод.
Действительно, в период 2,5-2,3 * 109 лет назад наблюдается резкое снижение числа строматолитовых формаций (Семихатов и др., 1999).
Разрушение архейских цианобактериальных сообществ дало возможность сложиться на их месте новым
(как обычно в таких случаях – более сложным и совершенным) сообществам.
В этих новых сообществах эукариоты уже являлись необходимым компонентом.
Первые эукариоты могли появиться задолго до конца архея, но только во время и после кризиса на рубеже архея и протерозоя
сообщества с их участием становятся заметным компонентом биоты,
и только с этого времени присутствие эукариот регистрируется в палеонтологической летописи.
(Марков А.В. Проблема происхождения эукариот)
Главная Науки о природе Биология :
Археозой: Об архее | О криптозое (2,5) | Катархей (4,6) | Эоархей (4,0) | Палеоархей (3,6) | Мезоархей (3,2) | Неоархей (2,8) | Галактические года
Ключевые слова для поиска сведений об археозойской и гадесской эрах:
На русском языке: неоархей, неоархейскская эра, второе глобальное окисление в археозое;
На английском языке: Neorchean.
|
|
|
|
|
|