|
|
Главная |
|
|
Пишите |
|
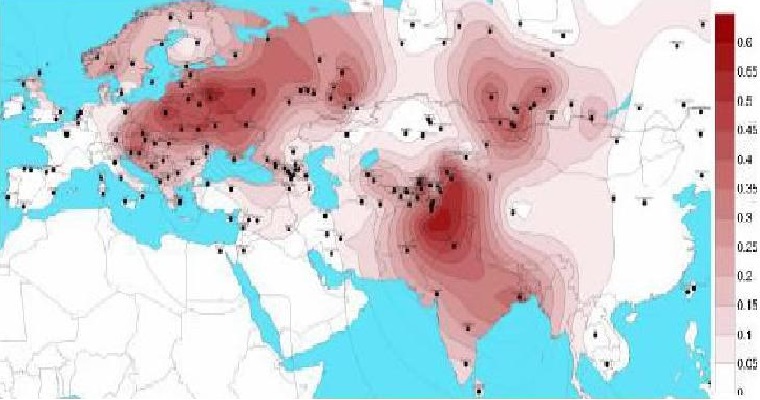
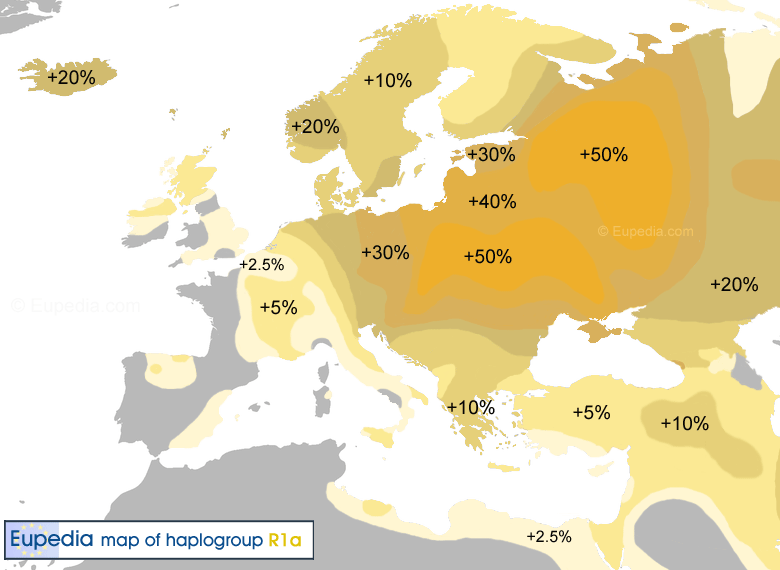
Гаплогруппа R1a появилась в эпоху верхнего палеолита предположительно в Южной Сибири примерно 20-15 тысяч лет назад, и на протяжении многих тысячелетий продвигалась на запад. Между 10 и 8 тыс лет R1a прибыли на Балканы. Ареал распространения гаппогруппы R1a от Исландии, Германии до Индии, Ирана и Алтая. Гаплогруппа R1a, как полагают, была доминирующей гаплогруппой среди северных и восточных индоевропейских народов, которые частью перешли в индо-иранскую (R1a1-Z93), частью в греческую (микенцы), фракийскую, балтийскую и славянскую ветви. Праиндоевропейцы [или их часть] возникли в ямной культуре (3300-2500 до н.э.), в Понтийской-Каспийской степи между современной Украиной и юго-западом России. Их расселение связано с одомашниванием лошадей в степях Евразии, и изобретением колесницы (см. выше R1b). В восточной части понтийско-прикаспийские степи в значительной степени связаны с индо-иранскими и балто-славянскими ветвями праиндоевропейского языка. На основании археологических, языковых и генетических данных, можно сказать, что скотоводы-кочевники, жившие в северных русских степях и лесостепях 7000 лет назад были преимущественно R1a по отцовскм линиям. В настоящее время, высокие частоты гаплогруппы R1a находятся в Польше (56% населения), Украине (от 50 до 65%), европейской части России (от 45 до 65%), Беларуси (45%), Словакии (40%), Латвии (40 %), Литве (38%), Чехии (34%), Венгрии (32%), Хорватии (29%), Норвегии (28%), Австрии (26%), Швеции (24%), северо-востоке Германии (23%) и Румынии (22%). Большинство носителей гаплогруппы R1a принадлежат к субкладу R1a1-M417 - его дерево смотрите в разделе ниже. А внутри этой гаплогруппы большинство принадлежит к популяции R1a1a1, которая зародилась на юге Восточной Европы 6 800 лет назад, и представители которой были носителями первых индоевропейских языков. На базе R1a1a1 сформировались славянские, индоарийские и другие этносы. |
Подгруппы гаплогруппы R1a: R1a* - R1a1 M17, M198 R1a1*- R1a1a M56 R1a1b M157 R1a1c M64.2, M87, M204
Разделы об Y-хромосомном субкладе R1a:
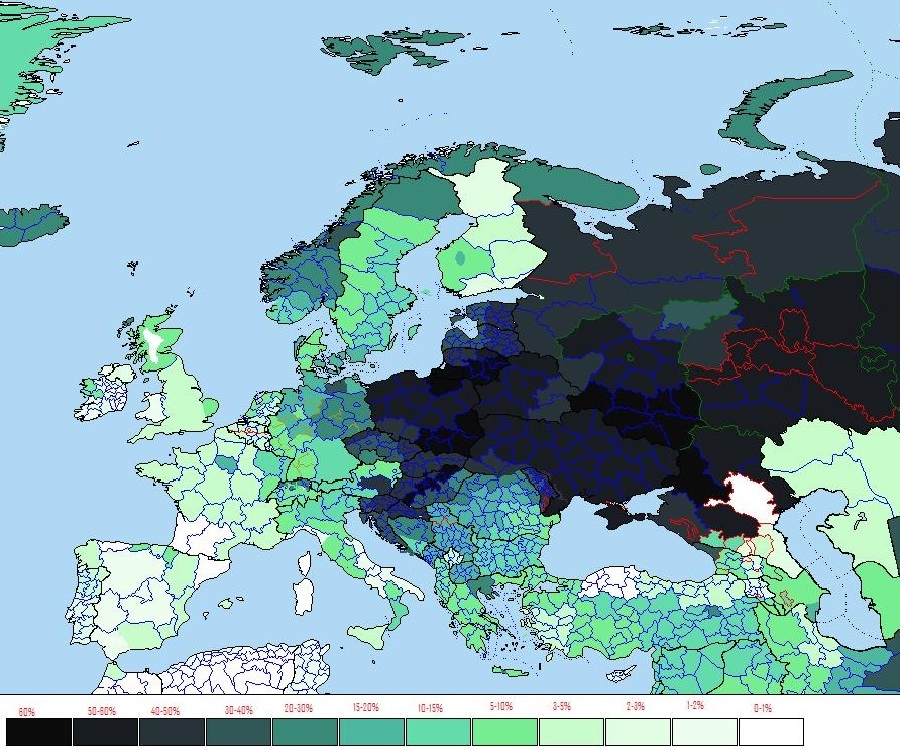
Наибольшее распространение имеет в Восточной Европе: среди лужичан (63 %), поляков (ок. 56 %), украинцев (ок. 54 %), белорусов (52 %), русских (47 %), татар 34 %, башкир (26 %) (у башкир Саратовской и Самарской обл. до 48 %); и в Центральной Азии: у худжандских таджиков (64 %), киргизов (63 %), ишкашими (68 %). Умеренное распространение в скандинавских странах (23 % в Исландии, 18-22 % в Швеции и Норвегии), в Иране ( 25 %). У брахманов индийских штатов Западная Бенгалия и Уттар-Прадеш гаплогруппа R1a1 встречается с частотой 72 % и 60 % соответственно (субклад характерный для Центральной и Южной Азии R1a1-Z93). Думаю, точнее связать гаплогруппу R1a не с именем славян, а с именем жителей Русской равнины, которыми тогда были, скорее всего, арийцы. Потом носителями ее стали скифы и сарматы - и только потом она перешла к славянам. Распространенность субкладов подтверждает это. Следовательно, славяне являются биологическими арийцами. В конечном же счете, и арийцы (R1a) и западноевропейцы (R1b) происходят от древних уральцев. |
Гаплогруппа R1a наиболее характерна для славян. Например, у русских распространены следующие гаплогруппы:
Здесь 3 самые распространённые гаплогруппы показывают какие этносы внесли свой вклад в формирование русского народа:
По другой версии, в русском этносе есть четыре основных гаплогруппы – R1a, I1, I2 и N1c1 (жители Южной Балтии), плюс еще десяток минорных, которых среди русских от нескольких процентов до долей процента.
В процентах указана доля R1a от общей численности этноса (славяне и индоиранцы):
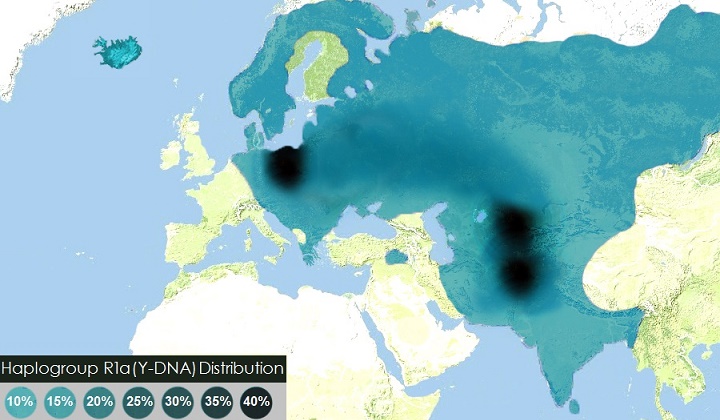
По современным данным, гаплогруппа R1a появилась в эпоху верхнего палеолита в Центральной Азии, предположительно в Алтайском регионе. Это соотносится с теми данными, что в том же регионе и в ту же эпоху обитали носители предковой гаплогруппы Р, от которой в результате серии SNP (Single Nucleotide Polymorphism), или снипов в Y-хромосоме, последовательно образовались гаплогруппы Q и R, далее R1, и далее R1a и R1b. Часть носителей гаплогруппы Q ушли через Берингов перешеек в Америку примерно 25-20 тысяч лет назад, прибыв туда по разным данным между 16 и 12 тысяч лет назад. Гаплогруппа R1a образовалась примерно 20 тысяч лет назад (у других исследователях - до 18-15 тысяч лет назад), и на протяжении многих тысячелетий продвигалась на запад, пройдя Тибет, Индостан (примерно 12 тысяч лет назад), Иранское плато, Анатолию (примерно 10 тысяч лет назад, где лингвистами впервые зафиксированы прото-индоевропейские языки между 9 и 7 тыс лет назад), и между 10 и 8 тыс лет назад прибыла на Балканы, неся свои протоиндоевропейские языки. Поэтому старый спор лингвистов, что является "прародиной" индоевропейских языков - Анатолия или Балканы, - получает определенный ответ ДНК-генеалогии - обе, в той же степени, что ни одна из них. Языки при длительных миграциях по континентам и на протяжении тысячелетий не имеют прародины, их развитие - длительный процесс конвергенции и дивергенции, и влияния многих прочих факторов. Согласно Андерхиллу с соавторами R1a появилась в восточной Европе около 9 300 до н.э., достигла территории Германии около 7 900 до н.э. и Дании около 7 700 до н.э. – Underhill et 2010). Распространение R1a в древних останках подтверждает предположение о распространении гаплоглогруппы R1a в центральную и северную Европу с востока. Носители гаплогруппы R1a обитали в Европе на протяжении нескольких тысячелетий, но распространение их (праиндоевропейского) языка в Европе на другие европейские гаплогруппы того времени - Е, F, G, I, J, K - остается неизвеcтным. Последние могли полностью или частично перейти на ИЕ языки, или сохранять свои древние языки. После прибытия в Европу эрбинов [?], носителей гаплогруппы R1b (4800-4500 лет назад для разных регионов Европы) почти все перечисленные выше гаплогруппы "старой Европы" или были истреблены, или частью бежали из Европы. Поэтому для перечисленных выше гаплогрупп современные их носители показывают "бутылочные горлышки" популяций в Европе, обычно в интервале 4800-3600 лет назад. Носители гаплогруппы R1a примерно 4600 лет назад передвинулись из Европы на Русскую Равнину (как культуры боевых топоров / шнуровой керамики, а также фатьяновская культура в понятиях археологии), и примерно 4500-4200 лет назад разделились на четыре арийские ДНК-генеалогические линии. Одна, потомки которой ныне представлены субкладом R1a-Z280 и дочерними субкладами, осталась на Русской равнине, и впоследствии частью мигрировала обратно на запад, в Европу, большей частью в ходе 1-го тыс до н.э. и первой половины 1-го тыс нашей эры. Три арийские линии, в основном субклада L342.2 и дочернего L657, мигрировали через Кавказ в Месопотамию (альтернативный вариант - из Средней Азии) и далее в Сирию, Ирак, Аравийский полуостров - это так называемые митаннийские арии; другое направление миграции - в Среднюю Азию и далее на Иранское плато (авестийские арии); третье направление - на Южный Урал и далее в Индию (индо-арии); все эти три основных направления миграции происходили в диапазоне 4200-3500 лет назад. Еще одна миграция ариев в те времена, которая фактически является продолжением миграции их на Южный Урал, продвинулась в Зауралье, образовав, например, карасукскую культуру (3800-3400 лет назад), и далее в Алтайский регион, на юг Сибири, в Монголию и далее до Северного Китая, внеся заметный вклад лексики, например, в древнекитайский язык. Часть из тех ариев влилась в состав тюркоязычных народов алтайского и северо-западного китайского региона. [Возможно, это были тохары, а не арии.] Учитывая интерес к ариям и вклад их в современную цивилизацию, гаплогруппа R1a получила название "арийской". В настоящее время до 72% состава высших каст в Индии имеют гаплогруппу R1a, которая особенно распространена среди браминов. Для сравнения, ни один из 367 тестированиях браминов не имел гаплогруппы R1b. Даже по одной этой причине гаплогруппа R1b не может рассматриваться как арийская. Более того, наука не имеет никаких данных, что до 1-го тыс до н.э. носители гаплогруппы R1b (эрбины) вообще говорили на индоевропейских языках. Как ни странно, это постулируется многими лингвистами при полном отсутствии каких-либо доказательств. Есть основания полагать, что ИЕ языки вошли в среду эрбинов только в начале-середине 1-го тыс до н.э. посредством быстрого распространения ИЕ языка кельтов, которые поначалу были носителями гаплогруппы R1a и прибыли с востока в ходе миграций носителей гаплогруппы R1a. В настоящее время гаплогруппа R1a среди этнических русских достигает 63%, в первую очередь на юге России. На севере она разбавляется гаплогруппой N1 (в первую очередь южно-балтийской гаплогруппой N1c1) и ее доля падает до 40-30% среди этнических русских. В Белоруссии, Украине, Польше для гаплогруппы R1a среди мужского населения достигает 50%. Высокое содержание гаплогруппы R1a-L342.2 имеет место среди таджиков, киргизов, узбеков, описаны ДНК-генеалогические линии R1a-L342.2 у башкиров, евреев. У всех этих народов общий предок линий R1a-L342.2 жил сравнительно недавно, от 2100+/-250 лет назад (киргизы) до 1300+/-150 лет назад (евреи) и до 1125+/-200 лет назад (башкиры). Субклад L342.2 указывает, что все эти линии имеют арийское происхождение, и появились в ходе миграций ариев через Месопотамию (евреи), Приуралье и Северный Казахстан (башкиры), Среднюю Азию (таджики и киргизы) во временном диапазоне 4500-3500 лет назад. |
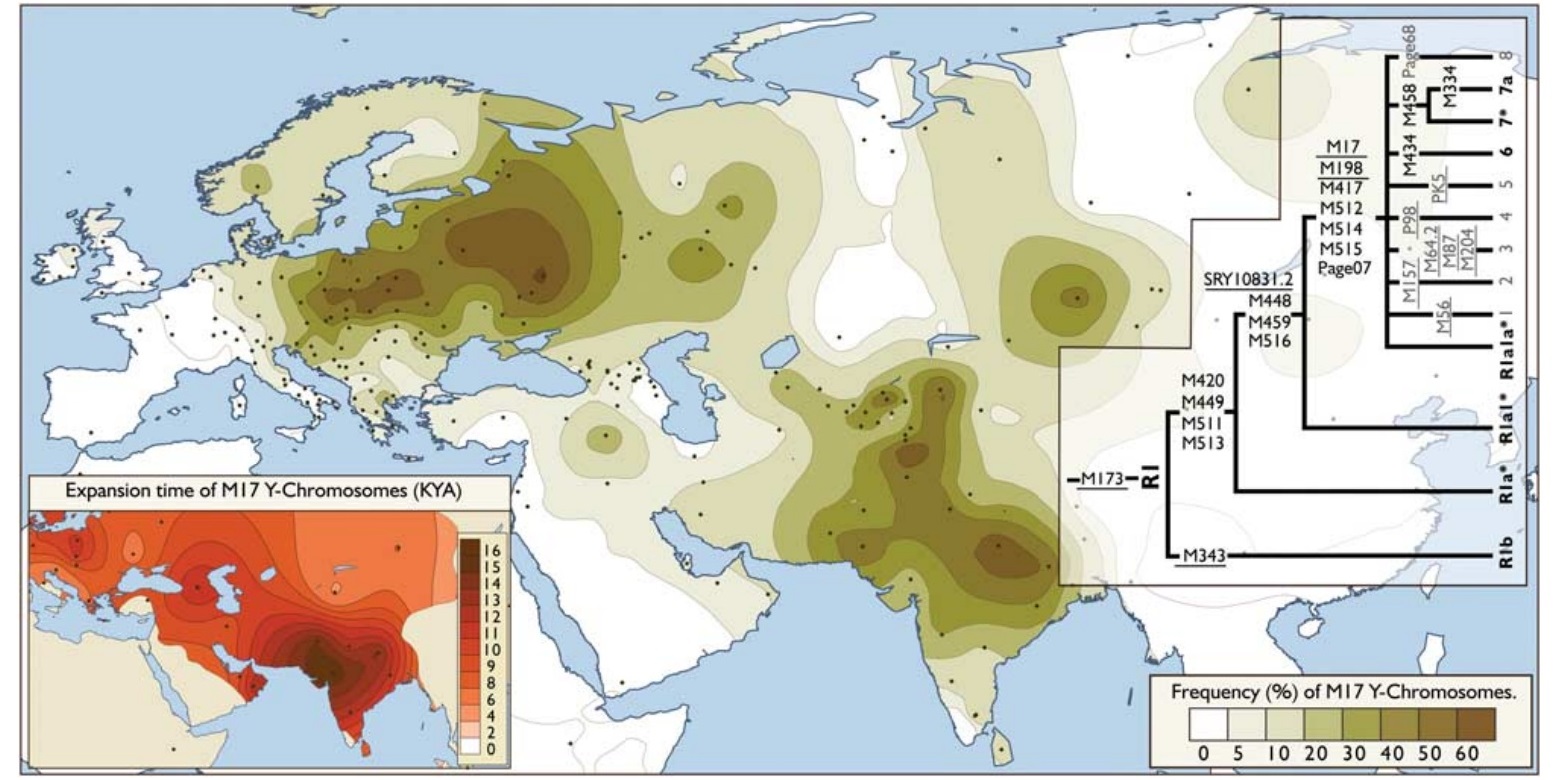
Гаплогруппа R1a1 ответвляется от гаплогруппы R1a во времена последнего ледникового максимума (вероятно около 19 000 лет назад), но точное место происхождения до сих пор неизвестно. Некоторые ученые предполагают, что, возможно, она возникла на Балканах или в районе Пакистана и Северо-Западной Индии, в связи с большим генетическим разнообразием, найденным в этих регионах. Тем не менее разнообразие можно объяснить и другими факторами. Балканы могли стать объектом миграций R1a1 около 7000 лет назад из евразийских степей. Южная Азия имела гораздо большую плотность населения, чем любой другой регион мира (иногда сопоставимо с Китаем), по крайней мере, 10 000 лет назад, и больше населения могло привести к большему генетическому разнообразию. Наиболее вероятным местом происхождения гаплогруппы R1a является Сибирь, а для гаплогруппы R1a1 - Центральная Азия или южная Россия. Гаплогруппа R1a1, как полагают, была доминирующей гаплогруппой среди северных и восточных прото-индо-европейских народов, которые эволюционировали в индо-иранскую, фракийскую, балтийскую и славянскую ветви. Индо-европейцы распространялись в ямную культуру (3500 до н.э.). Их расширение связано с быстрым распространением бронзового оружия, одомашниванием лошадей в степях Евразии и изобретением колесницы. В южной степной культуре, как полагают, имелись преимущественно R1b1 линии, а в северной лесостепной культуре была существенно доминирующей гаплогруппа R1a1. Первое распространение людей из лесостепи произошло с культурой шнуровой керамики. Миграции носителей R1b Центральной и Западной Европы способствовали распространению носителей гаплогруппы R1a1 в южной степи во времена катакомбной культуры (2800 лет до н.э.). Происхождение этой культуры очевидно связано с культурой шнуровой керамики и обильным использованием боевых топоров - две наиболее характерных особенности культуры шнуровой керамики. Этот процесс также мог начаться при сатемизации индо-европейских языков. Тестирование древних ДНК подтвердило наличие гаплогруппы R1a1 в останках из культуры шнуровой керамики в Германии (2600 лет до н.э.), у тохаров (2000 лет до н.э.) на северо-западе Китая, из погребений курганной культуры (около 1600 до н.э.) от андроновской культуры на юге России и на юге Сибири, а также из различных культур железного века на территории России, Сибири, Монголии и Центральной Азии. Исторически сложилось, что никакая другая часть Европы не была захвачена большее количество раз, чем степные Балканы. Хронологически первые захватчики R1a1 пришли на запад с расширением культуры шнуровой керамики (от около 3200 до н.э.), вторые - вторжение микенских греков (1600 до н.э.), затем фракийцев (1500 до н.э.), иллирийцев (около 1200 до н.э.), гуннов и алан (400 н.э.), авар, болгар и сербов (все около 600 н.э.), мадьяры N1c1 (900 н.э.). Эти народы вышли из разных мест евразийских степей, в регионе между Восточной Европой и Центральной Азией, что объясняет почему такие высокие разнообразие STR находится в пределах Балканского R1a1 в настоящее время. Это пока не представляется возможным определить этническое происхождение для каждой ветви гаплогруппы R1a1, кроме того, что R1a1 связаны со стпями Восточной Европы. |
В настоящее время, высокие частоты R1a1 находятся в европейской части России (от 45 до 65% населения), Польше (55%), Беларуси (49%), Украине (43%), Словакии (42%), Латвии (40%), Литве (38%), Чехии (34%), Венгрии (32%), Хорватии (29%), Норвегии (27%), Австрии (26%), на северо-востоке Германии (23%) и Швеции (19%).
Этот сценарий разрешает кажущееся противоречие между тем, что одни гаплотипы гаплогруппы R1a1 в Индии являются принадлежащими к субкладу Z93 (5500±500 лет), а другие индийские гаплотипы гаплогруппы R1a1 отличаются от них и по виду, и по возрасту и имеют общего предка примерно 7 тысяч лет назад. Это подавалось в статьях индийских и ряда европейских авторов как отрицание прихода ариев Z93 в Индию и как то, что все индийские гаплотипы гаплогруппы R1a1 являются автохтонными. И то, и другое скорее всего неверно. Приведём конкретные данные. В одной из работ приведены 46 шестимаркерных гаплотипов гаплогруппы R1a1 из трёх племён южной Индии, территория Андра Прадеш, племена Naikpod, Andh, и Pardhan. Все 46 гаплотипов имеют 126 мутаций 9000±950 лет до общего предка.
Рассмотрим серию пакистанских R1a1-Z93 гаплотипов, приведенных в той же статье Сенгупты. В ней - 42 гаплотипа, содержащих 166 мутаций, что даёт 9000±890 лет до общего предка. Эта величина в пределах погрешности совпадает с 9200±950, приведённой выше для южноиндийских древних гаплотипов. Возраст предка тоже совпадает в пределах погрешности расчётов (4375±800 и 4050±500 лет). Не считая «индоевропейского» предка, общий предок трёх пакистанских базовых гаплотипов гаплогруппы R1a1 жил не менее 12 тысяч лет назад. Это можно показать попарным сравнением базовых гаплотипов. Например, первый и третий гаплотип различаются на 9 мутаций на 10 маркерах. Это разводит их предков на 25 000 лет и помещает их общего предка на 15 000 лет назад. Второй и третий гаплотипы различаются на 8 мутаций на 10 маркерах. Это помещает их общего предка на 12 000 лет назад. В любом случае, это значительно древнее, чем гаплотипы R1a1 в Южной Индии. Видимо, это всё те же древние южносибирские гаплотипы, носители которых более 10 тысяч лет назад мигрировали в Китай, в Южную Индию, и в Европу. Предковый китайский гаплотип имеет характерную аллель DYS19=14, которая почти не встречается в Европе.
Наконец, кратко рассмотрим серию из 10 среднеазиатских гаплотипов гаплогруппы R1a1, приведённую в той же статье. Они содержат только 25 мутаций, что даёт 5000±900 лет до общего предка. Это та же величина, что и для «индоевропейских» индийских гаплотипов гаплогруппы R1a1. Таким образом, имеются два древних подразделения гаплотипов гаплогруппы R1a1 - одно на востоке и одно на западе. Восточные гаплотипы происходят от общих предков, живших 20-12 тысяч лет назад, и, видимо, мигрировавших южнее из региона южной Сибири, в Китай и далее в Южную Индию и Пакистан между 7 и 12 тысяч лет назад. Западные гаплотипы происходят в своём большинстве от общего предка, жившего около семи тысяч лет назад на Русской равнине, как будет показано в следующем разделе, но некоторые восходят к общему предку, жившему около 12 тысяч лет назад, видимо, в Европе. Около 4 тысяч лет назад имела место обратная миграция носителей гаплогруппы R1a1 с запада на восток, которая принесла гаплотипы с общим предком примерно 7 тысяч лет назад обратно в Сибирь, на территорию андроновской археологической культуры, в Среднюю Азию и далее в Индию, уже как «индоевропейские» гаплотипы гаплогруппы R1a1-Z93. Так в Индии встретились два потока гаплотипов гаплогруппы R1a1, имеющих разную историю, и различающихся по составу аллелей. Это и породило заблуждения в литературе в отношении отнесения этих гаплотипов, которые были ошибочно приняты за автохтонные индийские гаплотипы. Резюмируя этот материал, датировку и регион происхождения гаплогруппы R1a1 следует отнести к периоду примерно 20 тысяч лет назад, предположительно южная Сибирь. Вероятно, там же произошло образование гаплогруппы R1b, примерно 16 тысяч лет назад. Это все ветви древнего азиатского предка гаплогруппы R1а1. Примерно в то же время, около 12 тысяч лет назад, потомки азиатской ветви гаплогруппы R1a1 достигли Европы, и отсюда начинается история европейских ветвей гаплогруппы R1a1. Рассмотрим их историю, и, где возможно, географию.
Абсолютное большинство носителей гаплогруппы R1a1 принадлежат к субкладу R1a1a1 (R1a-M417) [или R1a1-M417?], который делится на следующие субклады:
Многие из Западной и Северной Европы R1a1, кто имеет отрицательный маркер Z284 подпадает под корень R1a1a1* (M417), или даже в старые R1a1a (M17) и R1a1 (SRY10831.2), которые происходят от древнейших известных R1a из лесостепи, культуры шнуровой керамики (см. ниже), которая предшествует либо всем вышеназванным субкладам, либо R1a1-Z283. Тем не менее, было обнаружено, что значительная доля R1a в Северо-Западной Европе, особенно в Норвегии, Англии, Ирландии и Исландии со значением 10 (вместо 12) на STR маркере DYS388. Среди них, некоторые люди были определены в качестве несущих мутацию L664. Происхождение старших субкладов (M17 и SRY10831.2) до сих пор неясно (возможно, мезолитические охотники-собиратели ранней Европы).
При изучении истории гаплогруппы R1a1, оказалось, что потомки старейших предков гаплогруппы R1a1 живут в настоящее время в Китае. Их общий предок жил примерно 20 тысяч лет тому назад, видимо, в Южной Сибири, где и образовалась также, возможно, гаплогруппа R1b. Более точных сведений пока не имеется. По другим данным это могло произойти в регионе Южной или Средней Азии. Как это удалось установить? В статье, озаглавленной «Физическая антропология и этничность в Азии: переход от антропометрии к геному», были приведены относительные частоты гаплогрупп, встречающихся у нескольких популяций Китая. Сами гаплотипы не сообщались, но было указано, что гаплогруппа R1a1 встречается у этнических групп Hui, Bolan, Dongxiang и Sala на севере Китая, с частотами 18, 25, 32 и 22%, соответственно, наряду с гаплогруппами DE, С, F*(xK), О, 03, К*(хО,Р) и P*(xR). Как видно, доля гаплогруппы R1a1 в этих популяциях весьма заметна причем именно древних и первых носителей гаплогруппы R1a1. Базовый гаплотип в столь древних гаплотипах определяется с трудом, и определённое количество мутаций может быть неточным, потому был применён пермутационный метод расчета времени жизни общего предка. Для этого метода не требуется базовый гаплотип, и не нужно вносить поправку на возвратные мутации. Для всех 31 гаплотипов сумма квадратов мутационных различий между всеми аллеями каждого маркера попарно была равна 10184. Её следует разделить на квадрат числа гаплотипов в серии (961), на число маркеров в гаплотипе (5) и на 2, поскольку пермутации считаются симметрично в обоих направлениях. Это дало среднее число мутаций на маркер 1.060, уже с учётом возвратных мутаций. Разделив на 0.00135 мут/маркер/поколение, получаем, что общий предок всех 31 гаплотипов жил 20 000 ±3000 лет назад. Это - более точная величина, чем рассчитанная выше, по «линейному» методу, хотя и совпадает с ней в пределах погрешности расчётов (21000±3000 лет до общего предка). Вполне вероятно, что гаплогруппа R1a1 (точнее, новая мутация М17) появилась примерно 20 тысяч лет назад в южной Сибири, и её носители разделились. Одни мигрировали на запад и дошли до Балкан примерно 12 тысяч лет назад, хотя этот общий предок технически мог относиться к любой части миграционной траектории. Другие носители гаплогруппы R1a1 Ушли южнее, в Китай и далее в южную Индию и Пакистан, куда прибыли примерно 12-7 тысяч лет назад, как будет показано ниже. Естественно, эти южно-индийские гаплотипы гаплогруппы R1a1 должны принципиально отличаться от «арийских», «индоевропейских» гаплотипов R1a1. Так оно и получилось.
Первое расширение R1a1 на запад состоялось с распространением культуры шнуровой керамики (или боевых топоров) 3200-1800 до н.э. с родины ямной культуры. Это была первая волна R1a1 в Европе, которая отвечает за наличие этой гаплогруппы в Скандинавии, Германии и части R1a в Чешской Республике, Словакии, Венгрии и Польше. Высокая распространенность R1a1 в балто-славянских странах в настоящее время является не только за счет расширения шнуровой керамики, а также длинного ряда позднейших миграций из России, последняя из которых состоялась с 5 по 1-й века нашей эры.
Германские ветви индоевропейских языков, вероятно, произошли от слияния R1a1 культуры шнуровой керамики (прото-славянский язык), и позднее прибывших итало-кельтских R1b из Центральной Европы. Это подтверждается тем фактом, что германский народ состоит как из R1a1, так и R1b1 (и эти две гаплогруппы пришли по отдельным маршрутам в разное время), а также по лингвистике прото-германских языков, который имеет сходство с итало-кельтскими и славянскими языками. [Это похоже на тохар, которые также являются носителями субкладов R1a и R1b, и их язык также близок как к кельто-италийским, так и балто-славянским. Может быть, тохары еще ближе к германским племенам?]
R1a1 Шнуровой керамики смешались с германскими аборигенами I1 и создали северные культуры бронзового века (1800-500 до н.э.). R1b предположительно достигли Скандинавии позже миграциями на север от современной гальштатской культуры (1200-500 до н.э.). Первый подлинный германский язык, по оценкам лингвистов, возник около (или после) 500 г. до н. Это подтверждает, что она превратилась в смесь Гальштатской культуры прото-кельтов и праславянской культуры шнуровой керамики. Уникальность некоторых из германских слов основана на заимствованиях из других языковых ветвей ИЕ языков. Кельтский язык, как известно, имеет много заимствованных слов из афрозийских языков, от ближневосточных мигрантов в Центральной Европе. Тот факт, что современная Скандинавия состоит из примерно 40% I1, 20% R1a и 40% R1b подтверждает идею о том, что германская этнической принадлежность и язык приобрел три-гибридный характер в железном веке.
Балтийские ветви, как полагают, произошли от фатьяновской культуры (3200-2300 до н.э.), северо-восточное расширение культуры шнуровой керамики (боевых топоров). В ранний бронзовый век R1a1-кочевники из северных степей и лесостепей смешиваются с коренным уральским населением (N1c1 линий) в данной области. Это подтверждает сильные позиции как R1a1, так и N1c1 гаплогрупп из южной Финляндии в Литве и прилегающей части России.
Происхождение праславян восходит приблизительно к 6 веку нашей эры. Славянские ветви дифференцированы, когда сама культура шнуровой керамики (см. германские ветви выше) поглощается Кукутень-трипольской культурой (5200-2600 до н.э.) из Западной Украины и северо-востоке Румынии, которая, кажется, была составлена в основном из I2a2 линий происходящих непосредственно из палеолита европейцев, с небольшой примесью ближневосточных иммигрантов (в частности, E-V13 и T). Таким образом, появились гибридные культуры шаровидных амфор (3400-2800 до н.э.) на территории нынешней Украины, Белоруссии и Польши. Распространение в этот период I2a2, E-V13 и T (наряду с R1a1) вокруг Польши, Беларусии и западной части России, объясняет, почему восточные и северные славяне (и литовцы) имеют значительную частоту гаплогруппы I2a2 с небольшим количеством E и Т. Через несколько веков, эта гибридизованная культура исчезла в доминирующей культуре шнуровой керамики.
Культура шнуровой керамики сопровождалась тержинецкой культурой (1700-1200 до н.э.), лужицкой (1300-500 до н.э.), чернолесской (1025-700 до н.э.) и милоградской (600 до н.э.-100 г. н.э.) культурами на северо-востоке от славян. Последняя важная славянская миграция, как полагают, произошла в 6-ом веке нашей эры, из Украины в Польшу, Чехию и Словакию, заполняя вакуум, оставленный от восточных германских племен, которые вторглись в Римскую империю [бред - типа, Польша была готской].
На прото-индо-иранском языке говорили люди, которые затем назвали себя «арийцами» в Ригведе и Авесте. Возникновение относится к синташтинской культуре (2100-1750 до н.э.), в Тобольской и Ишимской долинах, к востоку от Уральских гор, основанную скотоводческими племенами культуры Абашево (2500-1900 до н.э.), начиная с верхнего Дона, Волги до Урала, и культуры Полтавка (2700-2100 до н.э.), которая простирается от Нижнего Дона, Волги до Каспийской низменности. Синташтинско-Петровская культура была первым этапом прихода в Бронзовый век индо-европейцев к западу от Урала, открыв путь к обширным равнинам и пустыной Центральной Азии, богатой металлами в горах Алтая. Арийцы быстро распространилась по всей Центральной Азии, от берегов Каспийского моря до Южной Сибири и Тянь-Шаня, через торговлю, сезонные миграции стад, грабежи и набеги.
Упряжка лошадей и использование колесниц в военном деле, вероятно, были изобретены людьми синташтинской около 2100 до н.э. и быстро распространились в горной области Бактрии, Маргианы (современная граница Туркменистана, Узбекистана, Таджикистана и Афганистана). Медные рудники интенсивно использовались на Урале, и прото-индо-иранцы от синташтинско-петровской культуры экспортировали медь в огромных количествах на Ближний Восток. Вероятно, они также использовали природные ресурсы Зеравшанской долины, обустраивая медно-добывающии колонии около 1900 до н.э., также добывали олово. Олово было особенно ценным ресурсом в конце бронзового века, так как оружие из сплава меди и олова прочнее чем более примитивные из мышьяка и бронзы. В 1700-ых до н.э., индо-иранцы расширили границы до нижнего течения в долине Амударьи и занялись орошением в сельскохозяйственных общинах. К 1600 году до н.э., старые укрепленные города Маргианы и Бактрии были заброшены, затоплены северные степи. Группа культур Центральной Азии подверглась индо-иранскому влиянию известеному как андроновский горизонт, и этот процесс продолжался до 800 г. до н. э.
Индоиранские миграции продвинулись дальше на юг через Гиндукуш. К 1700 г. до н.э., верховая езда скотоводов проникла в Белуджистан (юго-запад Пакистана). Долину Инда уступили около 1500 до н.э., и северная и центральная части Индийского субконтинента были захвачены уже около 500 года до нашей эры. Миграции на запад распространили санскрит и боевые колесницы в Ассирию, где они стали популярны у правителей Митанни около 1500 до нашей эры. Мидии, парфяне и персы, все иранские докладчики из андроновской культуры, переехали на иранское плато около 800 до нашей эры. Те, кто остался в Центральной Азии остались в истории, как скифы, а потомки Ямной Культуры, которые остались в Понтийско-Каспийской степи стали известны как сарматы у древних греков и римлян.
Индоиранские миграции привели к высокой частоте гаплогруппы R1a на юге Средней Азии, в Иране и Индии. Наиболее высокая частота гаплогруппы R1a1 (около 65%) достигается в регионе вокруг Кыргызстана, Таджикистана и севере Афганистана. В Индии и Пакистане, R1a1 колеблется от 15 до 50% населения, в зависимости от региона, этнические группы и касты. R1a, как правило, сильнее распространены на северо-западе субконтинента, и слабее в дравидийских общинах на юге (Тамил-Наду, Керала, Карнатака, Андхра-Прадеш) из Бенгалии на восток. Около 50% брахманов (высшая каста в индуизме) принадлежат к гаплогруппе R1a1-L657, в связи с эффектом основателя.
Материнские линии в Южной Азии, тем не менее, в подавляющем большинстве не индоевропейские. Например, в Индии более 75% "родной" мтДНК M и R линий и 10%- из Восточной Азии. В остальных 15% случаях, примерно половина имеют происхождение из Среднего Востока. И только около 7 или 8% может быть условно "европейской" (из понтийско-каспийский степей) по происхождению, в основном в виде гаплогруппы U2 и W (хотя происхождение U2 по-прежнему под вопросом). Европейские линий мтДНК гораздо более распространены в Центральной Азии, и даже в Афганистане и на севере Пакистана. Это говорит о том, что индоевропейские вторжения в Индию были проведена в основном мужчинами путем войны, и первым крупным поселением женщин где распространены условно-европейские мито-гаплогруппы - это северный Пакистан, Западная Индия (Пенджаб в Гуджарате) и север Индии (штат Уттар-Прадеш), где гаплогруппы U2 и W являются наиболее распространенными.
В 1934 году шведский археолог Фольке Бергман обнаружил около 200 мумий светловолосых и европеоидных в Таримской впадине на северо-западе Китая (район, известный как Синьцзян, Восточный Туркестан или Уйгурия). Самые старые из мумий датируются 2000 г. до н.э. и 7 мужчин проверены учеными в 2010 г. и были положительны на SNP мутации, характерные для гаплогруппы R1a1. Современные жители бассейна реки Тарим - уйгуры, относятся как к гаплогруппе R1b-M73 (около 20%) и гаплогруппе R1a1 (около 30%).
Первые теории о происхождении таримских мумий гласят, что это группа ранних наездников из репинской культуры (3700-3300 до н.э.) мигрировали из Дон-Поволжья в горы Алтая, основав афанасьевскую культуру (ок. 3600-2400 до н.э.), откуда они двинулись на юг к Таримской впадине. Другая возможность состоит в том, что Таримские мумии происходят от прото-индо-иранского народа (см. выше), который расширил свои границы по всей Центральной Азии около 2000 года до нашей эры из синташтинско-петровской культуры. Отдельные группы перешли Тянь-Шань и в конечном итоге попали в Таримскую впадину. Эта теория имеет преимущество по датировкам Таримских мумий. В любом случае, большинство из мумий проверены в том числе и на мтДНК и установлено, что в основном они принадлежали к монголоидной гаплогруппе C4, и лишь несколько из них европейских и ближневосточных гаплогрупп (H, K и R).
Существует некоторая полемика в отношении возможной связи между Таримскими мумиями и тохарским языком, ветвью кентумного языка индо-европейской семьи, которые были распространены в Таримской впадине с 3 по 9 века нашей эры. Легко предположить, что Таримские мумии были от прото-тохарских миграций за счет соответствующего расположения и связей с индо-европейскими языками. Тем не менее, Таримские мумии предшествовали появлению тохарского языка более двух тысячелетий, и тохарский является кентумным языком, который не может быть потомком сатемных от прото-индо-иранской ветви. Другие ветви кентумных связаны с гаплогруппой R1b, а тохарский является единственным восточным кентумным языком, вполне возможно, что тохарский язык связан с Центрально-Азиатскими миграциями и субкладом R1b1-M73, а также с предками современных уйгуров, населявших бассейн Тарима.
Современные жители Центральной Азии, от Синьцзяна до Турции и от Волги до Гиндукуша, говорят в подавляющем большинстве на тюркских языках. Это может быть удивительно, так как это соответствует области, где индо-иранские ветви индо-европейских миграций были распространены, в бронзовом веке - андроновская культура, в железном веке - скифы. Так почему же индо-европейские языки на Востоке выживают только в славянской России, и в южной части Центральной Азии, в таких странах, как Таджикистан, Афганистан и некоторые части Туркменистана? Почему уйгуры, узбеки, казахи и кыргызы, а также современные степные народы понтийско-каспийского региона (крымские татары, ногайцы, башкиры и чуваши) говорят не на индоевропейском наречии? Генетически эти люди несут индоевропейские R1a1, и в меньшей степени, R1b линии. Это объясняется тем, что тюркские языки заменили иранские языки в Центральной Азии между 4-м и 11-м веками нашей эры.
Прото-тюркские языки возникли в Монголии и Южной Сибири и распространялись с кочевыми племенами гуннов. Они принадлежат к алтайской языковой семье, это монгольский и маньчжурский (некоторые из них также включают корейский и японский, хотя они имеют очень мало слов из общего словаря). Пока неизвестно, когда прото-тюркские впервые появилась, но их распространение началось с гуннскими миграциями на запад через евразийские степи, и всю Европу, но остановились на границах Римской империи. Гунны были потомками хунну. Древние тесты ДНК показали, что хунну были уже гибридные люди 2000 лет назад, со смешанной европейской и Северо-Восточно азиатскими линиями Y-ДНК и мтДНК. Современные жители этого региона имеют примерно 90% монгольских линий против 10% европейских. Самое старое присутствие европейских мтДНК по Монголии и Байкалу относится к более чем 6000 лет назад.
Похоже, что тюркский язык быстро заменил скифский и другие иранские диалекты по всей Центральной Азии. Другие миграционные волны принесли больше тюркских в Восточной и Центральной Европы, как и хазары, авары, болгары и турки (=> см. 5000 лет миграций из евразийских степей в Европу). Все они были на самом деле центрально-азиатскими кочевниками, которые приняли тюркский язык, но практически не имели никакой крови из Монголии. Тюркские вторжения поэтому внесли большой вклад больше в распространение индоевропейских линий (особенно R1a1) в Восточной Азии.
Тюркские языки не сохранились в Европе за пределами понтийско-каспийской степи [не так - в Молдавии живут гагаузы - потомки тюрков-печенегов]. Болгарский язык, несмотря на то, что назван в честь тюркского племени, на самом деле относиться к славянским языкам с мягким тюркским влиянием. Венгерский, иногда ошибочно принимают за наследника гуннского из-за названия, на самом деле это уральская группа языков (Magyar). Десятки тюркских языков в современном мире имеют высокую степень взаимопонимания в связи с их сравнительно недавним общим происхождением и кочевым характером (до недавнего времени). Две основные ветви можно рассматривать как два языка, также далеких, как испанский и итальянский языки с языками из других групп ИЕ.
Мало что известно о прибытии прото-греческого языка из степи. Микенская культура началась около 1650 г. до н.э. и, несомненно, является степной культурой. Тесная связь между микенским и прото-индо-иранскими языками предполагает, что они разделились довольно поздно, в период между 2500 и 2000 годах до нашей эры. Археологически, микенские колесницы, наконечники копий, кинжалы и другие бронзовые предметы демонстрируют поразительное сходство с Сейсмо-турбинской культурой (ок. 1900-1600 до н.э.) в северной русской лесостепи, известной большой подвижностью своих кочевых воинов (первоначально находились в Монголии [!]). Поэтому вероятно, что микенцы спустились из России в Грецию в период между 1900 и 1650 до н.э., где они смешались с местным населением, чтобы создать новый уникальный образ греческой культуры.
Если рассматривать распространенность рода R1a1a в мировом масштабе, то основной пик частот приходится на Восточную Европу. Наиболее высокие частоты гаплогруппы находятся в России, Украине и Польше. Два других ареала — это Южная Сибирь [бывшие иранцы] и Индостан [индоарийцы]. [Собственно, это поввторяет распределение R1a в целом.]
Гаплогруппа R1a1a1g1 встречается главным образом в Центральной и Восточной Европе. Основной ареал ограничен Польшой, Чехией и Словакией. В центральной и южной части Польши частота гаплогруппы достигает своего максимума - порядка 40%, в то время как на севере, востоке и юго-западе от 15 до 23%. В Чехии - 25%, в Словакии от 18 до 25% гаплогруппы R1a1a1g1.
Общий предок для этой линии именуемой 'Западные Анты' мог жить примерно 2500 лет назад. Найдена в основном у поляков.
Общий предок для этой линии именуемой 'Прусской' мог жить примерно 2500 лет назад. Встречается в основном у людей, чьи предки выходцы из Померании и Восточной Пруссии.
Ключевые слова для поиска сведений по гаплогруппе R1a Y-ДНК:
На русском языке: Y-хромосомная гаплогруппа R1a, арийская R1a-популяция;
На английском языке: Y-haplo-group R1a.
|
|
|
|
|
|